Bollettino di Produzione Integrata e Biologica
E' disponibile il Bollettino n.03 del 26 febbraio

IL LEGNO NERO DELLA VITE SI TRASMETTE POCO CON L'INNESTO
(pubblicato su L'Informatore Agrario del 2007, 53-58, n.40)
di R. Credi(1), F. Terlizzi(1), L. Milanesi(2), R. Bondavalli(3), F. Rizzoli(4), V. Vicchi(4)
(1)Dista (Dipartimento di scienzee tecnologie agroambientali) e Crive (Centro interdipartimentale per le ricerche viticole ed enologiche) Università di Bologna rcredi@agrsci.unibo.it
(2)Consorzio fitosanitario provinciale di Modena
(3)Consorzio fitosanitario provinciale di Reggio Emilia
(4)Servizio fitosanitario, Regione Emilia-Romagna, Bologna
Lavoro svolto nell'ambito del progetto «Studio sui giallumi da fitoplasmi della vite» finanziato dalla Regione Emilia-Romagna (lr 28/98 e lr 9/2000) tramite il Crpv (Centro ricerche produzioni vegetali).
In Europa le fitoplasmosi della vite più diffuse ed economicamente importanti sono due: il legno nero (LN) e la flavescenza dorata (FD).
I danni che causano possono essere ingenti perché le piante colpite deperiscono, talvolta muoiono, e in presenza di alte incidenze si registrano significative perdite di produzione a livello quanti-qualitativo.
Tali malattie, pur essendo molto diverse sotto l'aspetto eziologico ed epidemiologico, hanno la particolarità di manifestarsi con un'identica sintomatologia (Belli et al., 1994).
Per la loro precisa diagnosi si deve quindi ricorrere alle moderne metodologie analitiche di tipo biomolecolare. Mediante queste tecniche, quali la reazione a catena della polimerasi (PCR) e l'analisi del polimorfismo della lunghezza dei frammenti di restrizione (RFLP), i fitoplasmi associati al LN e alla FD sono stati anche geneticamente caratterizzati e diversamente classificati (Lee et al., 1998).
Negli ultimi tempi si sono purtroppo verificate gravi epidemie di FD in molti comprensori viticoli del Nord Italia.
La situazione che si è venuta a creare è apparsa talmente preoccupante che da parte del Ministero delle politiche agricole e forestali è stato emanato un apposito decreto di lotta obbligatoria.
Come in altre regioni, anche nelle più rinomate zone viticole dell'Emilia-Romagna si è proceduto agli opportuni controlli (Credi et al., 2001). Il fitoplasma della FD, che sembrava confinato solamente nelle province occidentali di Piacenza, Parma, Reggio Emilia e Modena, ora risulta, purtroppo, in fase di ulteriore espansione, cominciando a interessare anche alcuni comprensori delle province di Bologna e Ravenna. Il fitoplasma del LN è invece distribuito su tutto il territorio regionale. I primi scoppi epidemici di questa affezione comparvero all'inizio degli anni Ottanta con alte percentuali di viti colpite (Credi e Babini, 1984). La sua diffusione è stata poi generalmente limitata, ma ultimamente il LN si presenta di nuovo in fortissima crescita, configurandosi come una vera e propria emergenza fitosanitaria. Ciò è sempre più motivo di allarme e preoccupazione presso i vari operatori della filiera viti-vinicola.
Le caratteristiche epidemiologiche del LN sono molto complesse e ancora non completamente definite. È noto comunque che il suo agente causale, fitoplasma dello Stolbur, in natura viene principalmente veicolato dal cixiide Hyalesthes obsoletus, specie polifaga e solo frequentatrice occasionale della vite. Date le peculiarità di questo insetto, si deve ritenere che l'acquisizione e successiva trasmissione del fitoplasma non si verifichi da vite a vite. L'infezione viene acquisita invece da piante erbacee spontanee, ad esempio ortica comune (Urtica dioica) e vilucchio comune (Convolvulus arvensis), che svolgono il ruolo chiave di ospiti sia dell'insetto vettore sia dell'agente patogeno (Maixner et al., 1995; Alma e Conti, 2002; Credi et al., 2004).
Oltre alla suddetta modalità c'è poi la possibilità che il fitoplasma del LN venga trasmesso, e quindi diffuso, anche mediante l'uso di materiali di moltiplicazione infetti (Credi et al., 1990; Osler et al., 1997; Pavan et al., 1997; Osler et al., 2002). La conferma e la relativa quantificazione di questo aspetto sono molto importanti e soprattutto necessari per spiegare l'origine dell'alta incidenza della malattia osservabile sempre più frequentemente a danno dei giovani vigneti. Così, mediante la tecnica d'innesto che usualmente viene utilizzata nella pratica vivaistica, sono state intraprese delle specifiche prove di trasmissione. I risultati ottenuti con le nostre osservazioni vengono qui riportati e discussi.
Metodologia utilizzata nella sperimentazione
Nell'estate-autunno del 2003 e del 2004, nelle principali aree viticole delle province di Modena e di Reggio Emilia, si individuavano alcuni vigneti interessati dalla tipica sintomatologia associabile ai fitoplasmi.
Campioni fogliari di diverse viti ammalate venivano prelevati e sottoposti in laboratorio agli opportuni saggi diagnostici; i risultati analitici dimostravano poi la sola presenza del fitoplasma del LN. Gli impianti prescelti erano costituiti dai seguenti vitigni: Ancellotta, Lambrusco Salamino, Lambrusco Grasparossa e Lambrusco Marani, tutti molto suscettibili al fenomeno fitopatologico in studio. Le viti, di 6-10 anni d'età, risultavano allevate e potate con il sistema Sylvoz. L'incidenza di quelle sintomatiche, nei due anni considerati, variava dal 6% al 23%. Il livello di gravità dell'affezione veniva inoltre stimato in base alla sua distribuzione nell'ospite: sintomi localizzati (L) se le manifestazioni patologiche interessavano solo alcuni tralci (≤ 5), sintomi generalizzati (G) se interessavano invece più o meno tutta la vegetazione. Nelle seguenti stagioni invernali del 2004 e 2005 si procedeva a raccogliere e conservare i tralci di potatura da singole viti, sia ammalate sia asintomatiche (A). Queste ultime erano da ritenersi confronti in apparenza sani e/o eventualmente con una qualche infezione di tipo latente (Osler et al., 2002). Nell'aprile 2004 e 2005 singole gemme, quali marze autoindicatrici dell'eventuale presenza del fitoplasma del LN nel materiale propagativo, venivano innestate su talee certificate di un comune soggetto (1103 P e SO4).
Relativamente alle prove del 1° anno, dal legno di alcune delle viti donatrici si preparavano anche delle talee. Parte di queste si sottoponevano a sola radicazione (franche) mentre altre, per riprodurre il ruolo di un soggetto eventualmente infetto dall'agente patogeno, erano invece innestate con gemme di un clone sano di Sangiovese (SG 12T). Questo vitigno risulta pure molto sensibile al LN (Belli et al., 1994).
La procedura impiegata era quella dell'innesto al tavolo mediante apposita macchina pneumatica a «omega», seguita da paraffinatura, forzatura, nuova paraffinatura e ambientamento. Le piante bimembri e quelle franche di piede venivano poi poste in vivaio e allevate adottando le usuali tecniche agronomiche, incluso periodici trattamenti insetticidi (foto A). Le barbatelle ottenute con le prove del 2004, durante l'inverno successivo sono state poste in conservazione e ripiantate nella primavera del 2005 per un'ulteriore valutazione.
In modo analogo si è proceduto per quelle del 2005, piantandole nuovamente nel 2006. A fine settembre dei bienni 2004-05 e 2005-06, le singole giovani viti erano oggetto di attente osservazioni sintomatologiche; quelle giudicate ammalate da LN e/o con delle manifestazioni dubbie venivano contrassegnate, campionate e poi analizzate mediante la tecnica PCR. Tutte le attività per la realizzazione-conduzione delle prove sono avvenute presso le strutture tecniche di un'azienda vivaistica con sede a Brisighella (Ravenna).
Per i saggi diagnostici si procedeva all'estrazione del DNA dai campioni e a una sua prima amplifi cazione con una coppia di iniziatori universali (P1/P7), complementari cioè a un frammento di DNA di tutti i fi toplasmi. I prodotti della reazione erano poi sottoposti a una seconda PCR (nested), al fi ne di identifi care quelli associati al LN e alla FD. Allo scopo si impiegavano le seguenti coppie di iniziatori: fStol/rStol, specifici per il fitoplasma del LN o del sottogruppo A del gruppo tassonomico dello Stolbur (16SrXII-A); R16(V)F1/R1, specifici per i fitoplasmi del gruppo del giallume dell'olmo (16SrV), fra i quali viene classificato quello della FD.
Tali coppie di iniziatori amplificano rispettivamente frammenti di DNA di circa 570 e 1.100 paia di basi. La dimostrazione di ciò avveniva sottoponendo il prodotto a elettroforesi in gel di agarosio e, previa colorazione con bromuro di etidio, visualizzando sotto luce ultravioletta le eventuali bande per le opportune valutazioni (Lee et al., 1998; Maixner et al., 1995; Credi et al., 2001; Credi et al., 2004).
Risultati
Considerando le prove di trasmissione effettuate nel 2004, la tabella 1 riassume i risultati degli innesti su 1103 P con le gemme-marze provenienti dal legno di potatura di 30 viti donatrici. Le barbatelle ottenute dai 1.772 innesti-talea messi a dimora sono state purtroppo solo 221. Il bassissimo attecchimento era dovuto alle avverse condizioni pedo-climatiche sopraggiunte durante il primo periodo di allevamento in vivaio. Fra le piante sopravvissute, 3 evidenziavano un anormale arrossamento internervale delle foglie, particolarmente a carico di quelle più vecchie, e una non perfetta lignificazione dei rispettivi tralci. Ritenendo però la manifestazione non proprio caratteristica del LN, tali barbatelle erano classificate come sospette (foto 2).
L'incertezza veniva poi delucidata mediante le analisi PCR che risultavano tutte negative (dati non presentati). Anche i rilievi effettuati il 2° anno dall'innesto non portavano alla sicura identificazione di viti ammalate da LN; infatti fra le 155 ripiantate solamente 3 piante mostravano sintomi di dubbia tipologia e nuovamente risultavano negative ai successivi test diagnostici.
Riguardo alle talee franche ottenute dal legno di 15 viti donatrici e quelle innestate con il clone sano di Sangiovese (SG 12T) derivate dai tralci di 16 viti donatrici, rispettivamente 169 e 161, solo una barbatella della 2ª prova manifestava i sintomi tipici di LN (tabella 2). Tale pianta risultava originata da una talea-soggetto proveniente da vite donatrice di Lambrusco Salamino e valutata ammalata con una distribuzione localizzata (L) l'autunno precedente. Le anomalie osservate consistevano principalmente in accentuato nanismo, ingiallimento-arrossamento più o meno settoriale delle lamine fogliari, anche a danno di quelle più giovani della cima, e mancata lignificazione del tralcio principale (foto 3). L'infezione dello specifico fitoplasma nella barbatella è stata poi confermata tramite PCR (dati non presentati). Per contro, mediante i rilievi effettuati il 2° anno sulle giovani viti ripiantate, 126 e 121 rispettivamente, non veniva individuato nessun caso con sospetta infezione dell'agente della malattia.
I dati ottenuti con la sperimentazione del 2005 (innesti su SO4 di gemme marze provenienti da 41 viti donatrici) sono sintetizzati nella tabella 3. Gli innesti-talea eseguiti sono stati complessivamente 2.309, ma quelli realmente attecchiti solo 1.085. Le osservazioni effettuate nel biennio non hanno individuato con certezza la presenza di barbatelle affette da LN. Sintomatologie dubbie venivano comunque registrate sia nel 1° anno di osservazione (5 barbatelle) che nel 2° (1 sulle 783 ripiantate); tuttavia in nessuna di queste i saggi PCR di verifica dimostravano la presenza di fitoplasmi.
Discussione
L'avvenuta trasmissione del fitoplasma del LN è stata da noi dimostrata solamente su una barbatella nell'ambito delle prove effettuate nel 2004. La pianta infetta veniva identificata fra talee-soggetti di viti originariamente ammalate di Lambrusco Salamino, innestate con gemme marze di un clone sano di Sangiovese. Se si considera il singolo gruppo nell'ambito del quale è stata registrata (tabella 2), la trasmissione dell'agente patogeno è risultata del 1,7%; facendo invece una valutazione media fra tutte le barbatelle ottenute innestando le talee delle varie viti donatrici, l'incidenza appare ovviamente molto più bassa (0,6%). Per contro, nessuna evidenza di riproduzione della malattia si è avuta nei gruppi delle piante franche di piede e sempre preparate con il legno di potatura sia di viti asintomatiche sia affette dal LN.
Stesso comportamento per quanto riguarda gli innesti effettuati su 1103 P con le gemme-marze provenienti dalle varie viti donatrici (tabella 1). I rilievi effettuati nel biennio non hanno infatti evidenziato casi certi che indicassero una trasmissione dell'affezione attraverso l'innesto; solo alcune giovani viti mostravano un tipo di sintomatologia giudicata poco specifica e con esito negativo ai saggi diagnostici di verifica. In riferimento a queste prove, occorre comunque aggiungere che lo scarso attecchimento delle talee, e più in particolare la bassissima percentuale (12,6%) di riuscita degli innesti su 1103 P potrebbero avere influenzato i risultati, apparendo cioè sottostimata la trasmissibilità di LN mediante i materiali propagativi utilizzati.
Tale considerazione non dovrebbe essere riferita alla prova effettuata nel 2005, dove la percentuale media di attecchimento degli innesti (47%) può considerarsi più accettabile sotto l'aspetto vivaistico.
Comunque, anche in questo caso, non si dimostra la trasmissione del fitoplasma del LN mediante la propagazione vegetativa. Con i rilievi svolti nel biennio, infatti, solo alcune piante con manifestazioni poco specifiche venivano osservate, risultando però tutte negative ai successivi controlli analitici (tabella 3).
In riferimento a ciò, è utile ricordare che in vivaio, anche a un occhio abbastanza esperto, altre anormalità di svariata natura potrebbero essere confuse con le ampelopatie da fitoplasma. Fra queste, si segnala l'accartocciamento fogliare, che come noto è indotto da diverse entità virali appartenenti ai generi ampelovirus e closterovirus (Martelli, 2002).
Dai dati scaturiti dalla realtà operativa che ha caratterizzato la nostra sperimentazione, emerge la scarsissima possibilità di trasmissione mediante innesto del fitoplasma di LN; questo considerando che sono stati utilizzati materiali di moltiplicazione provenienti da viti palesemente ammalate nel precedente periodo autunnale, alcune delle quali a quel tempo pure controllate mediante PCR e risultate positive alla presenza del fitoplasma.
La non registrazione di barbatelle ammalate fra quelle ottenute con il legno di viti donatrici asintomatiche, ma campionate negli stessi vigneti ove era presente il LN, escluderebbe, fra queste, l'esistenza di infezioni latenti. Tale particolarità risultava invece evidenziata dal lavoro di Osler et al. (2002) che, nel caso di marze di Chardonnay prelevate da piante asintomatiche ma presenti in vigneti con bassa incidenza di LN, ottenevano una trasmissione della malattia dello 0,1%. Per contro, la percentuale di trasmissione era nettamente superiore (3%) se le marze provenivano da viti sintomatiche.
Anche una precedente indagine svolta in Emilia-Romagna ha evidenziato che talee affette di Pinot bianco e Sangiovese, innestate con gemme sane di Baco 22A e Chardonnay quali marze indicatrici, rivelavano una percentuale media di trasmissione di LN rispettivamente del 4 e del 5,6%. Innestando invece talee sane di Kober 5BB con gemme di viti Chardonnay e Sangiovese variamente ammalate quali marze autoindicatrici, le incidenze medie delle barbatelle ammalate risultavano invece rispettivamente del 8,9 e del 2,6% (Credi et al., 1990).
La discordanza con i presenti risultati potrebbe derivare da un diverso comportamento, quale la superiore sensibilità all'infezione, dei suddetti vitigni (Belli et al., 1994). Inoltre, anche in base a quanto precedentemente accennato riguardo eventuali manifestazioni patologiche che possono ingenerare incertezze diagnostiche, è possibile che quelle percentuali di trasmissione fossero sovrastimate. A quel tempo, infatti, l'identificazione di LN e FD avveniva esclusivamente su base sintomatologica, senza cioè il supporto delle attuali metodologie molecolari.
Conclusioni
I risultati presentati sono un'ulteriore conferma della scarsissima importanza epidemiologica dei materiali di moltiplicazione vegetativa nella trasmissione dell'agente del LN alla progenie. A commento di ciò, e allo stato attuale delle conoscenze, si possono avanzare alcune ipotesi. Ad esempio, la scarsissima sistemicità del fitoplasma nell'ospite e, in particolare, negli organi legnosi quali i tralci. Tale caratteristica sarebbe suffragata dai risultati analitici accumulati nell'ambito di una specifica indagine sulla distribuzione dei fitoplasmi nelle piante colpite e dalla constatata alta frequenza del fenomeno del risanamento naturale fra le popolazioni dei vitigni considerati (dati non pubblicati).
La mobilità e la vitalità dell'agente patogeno potrebbero venire fortemente compromesse a seguito dei processi istopatologici che insorgono con l'infezione del microrganismo. I componenti del tessuto conduttore floematico a livello fogliare si presentano infatti estremamente disorganizzati, necrotici e/o schiacciati con parete ingrossata e collassata; nei tubi cribrosi apparentemente normali, si formano, invece, abnormi ammassi di callosio, osservabili specialmente a livello delle placche cribrose e che verosimilmente anche in questo caso ne impediscono la funzionalità (Credi, 1994).
Molto rimane da approfondire sull'argomento trattato. Un aspetto tuttora quasi completamente sconosciuto è il preciso ruolo svolto dagli ibridi portinnesti che, occorre ricordarlo, usualmente si comportano come portatori latenti delle infezioni fi toplasmatiche (Belli et al., 1994). Al proposito si richiama l'attenzione sul fatto che l'unico caso certo di trasmissione del LN da noi accertato è avvenuto proprio con una talea di vite donatrice impiegata come ipobionte. Con lo studio condotto pensiamo tuttavia di avere contribuito all'ampliamento delle conoscenze sulle caratteristiche epidemiologiche del LN. Le alte incidenze della malattia osservabili in giovani impianti dei vitigni presi in esame sarebbero quindi imputabili, nella quasi totalità, a infezioni naturali del fitoplasma da parte degli insetti vettori. Infezioni che in parte potrebbero anche verificarsi durante il periodo di crescita in vivaio delle barbatelle.
Seppure tale eventualità non risulti al momento sufficientemente indagata e quantificata, in tali siti è comunque buona norma profilattica monitorare e controllare le popolazioni di quelle piante erbacee spontanee sorgenti d'inoculo, che fungono cioè sia da ospiti dell'agente patogeno sia dell'insetto vettore Hyalesthes obsoletus (Maixner et al., 1995; Alma e Conti, 2002; Credi et al., 2004).
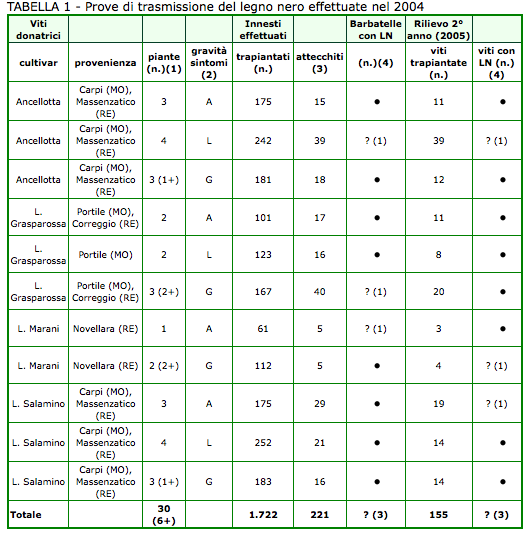
Innesti su 1103 P con gemme (marze autoindicatrici) prelevate da viti ammalate e asintomatiche di Ancellotta, Lambrusco Grasparossa e Lambrusco Salamino. (1) Fra parentesi numero di viti analizzate mediante PCR nell'autunno precedente e risultate positive (+) al fitoplasma del LN. (2) Valutazione della malattia nell'autunno precedente: A = viti asintomatiche, L = viti con sintomi localizzati, G = viti con sintomi generalizzati. (3) Riuscita degli innesti compromessa da sopraggiunte avverse condizioni pedoclimatiche. (4) - = nessuna pianta con sintomi evidenti; ? = piante con sintomatologia incerta (fra parentesi il numero), risultate poi negative all'analisi PCR.
Fra le piante sopravvissute, 3 ogni anno, pur evidenziando un anormale arrossamento internervale delle foglie e una non perfetta lignificazione dei tralci, mediante le analisi PCR risultavano tutte negative.
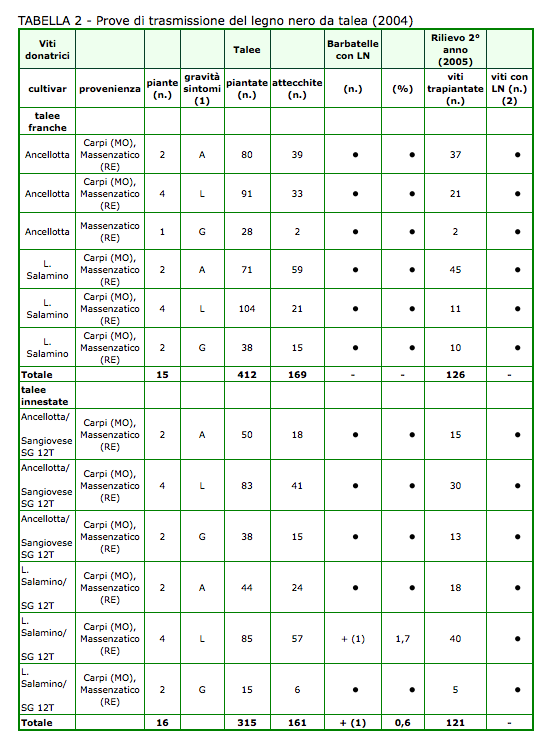
Talee prelevate da viti ammalate e asintomatiche di Ancellotta e Lambrusco Salamino, non innestate (franche) e innestate con gemme di un clone sano di Sangiovese (marze indicatrici). (1) Valutazione della malattia nell'autunno precedente: A = viti asintomatiche, L = viti con sintomi localizzati, G = viti con sintomi generalizzati. (2) - = nessuna pianta con sintomatologia evidente; + = piante sintomatiche (fra parentesi il numero) e infezione del fitoplasma confermata poi con analisi PCR.
Solo una barbatella della 2a prova manifestava i sintomi tipici del LN; l'infezione dello specifico fitoplasma veniva inoltre confermata tramite PCR. Nessun caso di sospetta infezione veniva invece rilevato l'anno seguente.
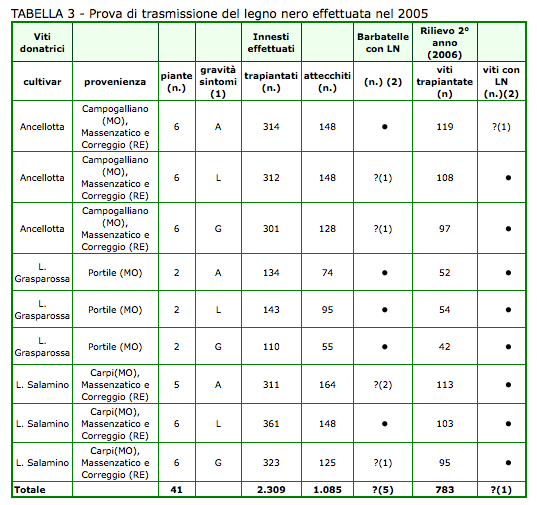
Innesti su SO4 con gemme (marze autoindicatrici) prelevate da viti ammalate e asintomatiche di Ancellotta, Lambrusco Grasparossa e Lambrusco Salamino. (1) Valutazione della malattia nell'autunno precedente: A = viti asintomatiche, L = viti con sintomi localizzati, G = viti con sintomi generalizzati. (2) - = nessuna pianta con sintomi evidenti; ? = piante con sintomatologia incerta (fra parentesi il numero), risultate poi negative all'analisi PCR.
Sintomatologie dubbie venivano registrate sia nel 1° anno di osservazione (5 barbatelle) sia nel 2° (1 sulle 783 ripiantate); tuttavia in nessuna di queste i saggi PCR di verifica dimostravano la presenza di fitoplasmi.
Per consultare la bibliografia
e gli approfondimenti di questo articolo visitare il sito: www.informatoreagrario.it/ita/Riviste/Infoagri/Lia4007/2701_web.pdf
BIBLIOGRAFIA
Alma A., Conti M. (2002) - Flavescenza dorata e altre fitoplasmosi della vite: il punto su vettori ed epidemiologia. Informatore fitopatologico, 10: 31-35.
Belli G., Credi R., Refatti E. (1994) - Recenti sviluppi nelle conoscenze sulla flavescenza dorata e altri giallumi della vite. Atti Giornate Fitopatologiche, 2: 295-306.
Credi R., Babini A.R. (1984) - Casi epidemici di giallume della vite in Emilia-Romagna. Vignevini, 3: 35-39.
Credi R., Santucci A., Martini L. (1990) - Trials on graft transmission of a grapevine flavescence dorèe-like disease. Phytopath. Medit., 29: 7-13.
Credi R. (1994) - Occurrence of anomalous mycoplasma-like organisms in grapevine yellow-diseased phloem. J. Phytopathology, 142: 310-316.
Credi R., Terlizzi F., Bissani R., Poggi Pollini C. (2001) - Presenza e diffusione dei fitoplasmi del legno nero e della flacescenza dorata della vite in Emilia-Romagna. Vignevini, 12: 107-110.
Credi R., Terlizzi F., Cricca L., Dradi D. (2004) - Epidemiologia del legno nero della vite. L'Informatore Agrario, 7: 72-75.
Lee I.M., Gundersen D.E., Davis R.E., Bartoszyk I.M. (1998) - Revised classification scheme of phytoplasmas based on RFLP analyses of 16S rRNA and ribosomal protein gene sequences. International Journal of Systematic Bacteriology, 48: 1153-1169.
Martelli G.P. (2002) - Le principali virosi della vite oggi. Informatore Fitopatologico, 4: 18-27.
Maixner M., Ahrens U., Seemuller E. (1995) - Detection of the German grapevine yellows (Vergilbungskrankheit) MLO in grapevine, alternative hosts and vector by a specifi c PCR procedure. European Journal of Plant Pathology, 1001: 241-250.
Osler, R., Vindimian M.E., Filippi M., Carraro L., Refatti E. (1997) - Possibilità di propagazione del giallume della vite (legno nero) a mezzo del materiale vivaistico. Informatore Fitopatologico, 11: 61-63.
Osler R., Zucchetto C., Carraio L., Frausin, C. Pavan, F., Vettorello G., Girolami V. (2002) - Trasmissione di fl acescenza dorata e legno nero e comportamento delle viti infette. L'Informatore Agrario, 19: 61-65.
Pavan F., Villani A., Fornasier F., Girolami V. (1997) - Ruolo del vivaismo nella diffusione della flavescenza dorata. L'Informatore Agrario, 10: 69-71.
ABSTRACT
Trials on graft transmission of grapevine bois noir phytoplasma
Trials using bench grafting to transmit grapevine bois noir (BN) phytoplasma were carried out in the Emilia-Romagna region of northern Italy. For indexing, plant leaf samples were tested by PCR using the P1/P7 oligonucleotide universal primers. A second pair of nested primers, fStol/rStol, were subsequently used to amplify a specific target sequence from the Stolbur phytoplasma. Healthy single-bud scions of a Sangiovese clone were grafted onto donor stock cuttings taken from diseased Ancellotta and Lambrusco Salamino vines. Afterwards, grafts were planted in the nursery and observed for a two-year period. Positive transmission was revealed during the autumn only in a Sangiovese indicator vine, out of 161, with an average transmission rate of 0,6%. Healthy 1103 P and SO4 rootstoks were also grafted with bud scions taken from Ancellotta, Lambrusco Salamino and Lambrusco Grasparossa BN-aff ected vines. Among 1.300 young vines, none displayed the typical disease symptoms. The epidemiological significance of these results is discussed.

E' disponibile il Bollettino n.03 del 26 febbraio

I Giallumi della vite (dall'inglese Grapevine Yellows, GY) sono malattie dovute a fitoplasmi che si …

Attraverso il decreto 21 Novembre 2016 viene concessa una proroga di 90 giorni sia per la commerc…

I nostri sforzi per diminuire l’impiego di insetticidi nei vigneti hanno fino ad oggi coinvolto se…

ARTICOLI CORRELATI Cocciniglie farinose


Il controllo del territorio e la gestione della problematica

PARTE GENERALE Da diversi anni in alcune aziende della provincia di Reggio Emilia si stanno rilevan…
Consorzio Fitosanitario Provinciale di Reggio Emilia - Via Gualerzi, 32 - 42124 - Reggio Emilia (RE)
Tel. 0522-271380 - Fax. 0522-277968 - mail: fitosanreggio@regione.emilia-romagna.it - Privacy policy
C.F.: 80011830355
